Tous droits réservés © NeurOreille (loi sur la propriété intellectuelle 85-660 du 3 juillet 1985). Ce produit ne peut être copié ou utilisé dans un but lucratif.
 Français
Français
 English
English
 Español
Español
 Português
Português
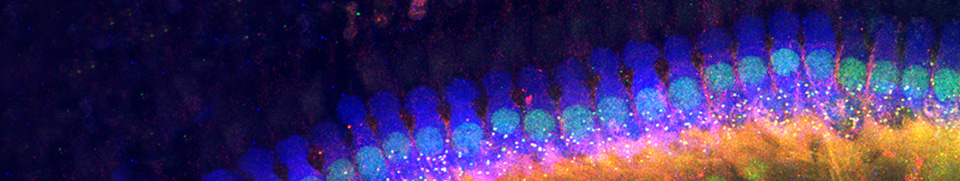
Os canais do labirinto membranoso coclear conteém dois tipos de fluidos: perilinfa e endolinfa. A perilinfa, com composição semelhante ao líquido extracelular, enche as rampas timpânica e vestibular. A endolinfa, que enche o canal coclear, tem uma composição única no organismo.
Composição dos fluidos cocleares
A composição da endolinfa é uma particularidade importante da cóclea. Com efeito, este líquido que enche a rampa média, é muito rico em potássio (150mM), muito pobre em sódio (1mM) e quase sem cálcio (20 a 30 µM).

A perilinfa (em azul) preenche as rampas vestibular (1) e timpânica (2).
A endolinfa (em verde) é o líquido da rampa média ou canal coclear(3), sendo muito rica em potássio segregado pela estria vascular e possuindo um potencial positivo de +80mV em relação à perilinfa.
Note que apenas a superfície do órgão espiral de Corti (designadamente os cílios das células ciliadas) é banhada pela endolinfa.
Perilinfa
Existem dois tipos de perilinfa: a da rampa vestibular e a da rampa timpânica; as duas têm uma composição muito semelhante à do líquido cefaloraquidiano: Rica em sódio (140 mM) e pobre em potássio (5mM) e cálcio (1,2 mM). a origem da perilinfa da rampa vestibular é diferente da da rampa timpânica. Enquanto que a perilinfa da rampa vestibular provém do plasma através da barreira hematoperilinfática, a perilinfa da rampa timpânica provém do líquido cefaloraquidiano.
Endolinfa
A endolinfa é, ela própria, gerada a partir da perilinfa.
O potencial endococlear é a soma de dois potenciais: um potencial positivo devido à secreção ativa de K+ pela estria vascular (+ 120mV), e um potencial negativo gerado pela difusão passiva de iões K+ pelas células ciliadas (-40mV). Este potencial pode ser posto em evidência após uma anóxia.
É de notar que o desenvolvimento da composição iónica da endolinfa precede o desenvolvimento do potencial endococlear. Com efeito, as concentrações definitivas de iões da endolinfa surgem durante a primeira de vida pósnatal no rato, enquanto que o potencial endococlear só se desenvolve a partir do 12º dia de vida pósnatal e só atinge os valores definitivos na 3ª semana de vida pósnatal.

Composição e propriedades dos dois fluidos cocleares
Porquê a endolinfa?
Parece que a cóclea encontrou um processo de fazer entrar e sair potássio sem dependência de ATP.
Em regra geral, se um ião entra numa célula de forma passiva, ele sairá de forma ativa e vice versa.
Só o polo apical das células ciliadas está banhado pela endolinfa, muito rica em K+ e que apresenta um potencial positivo de 80mV. Os iões K+ vão assim entrar de forma passiva nestas células visto haver uma maior concentração de K+ na endolinfa que no interior das células ciliadas e o seu potencial de repouso ser de - 60mV, o que favoriza a entrada de K+. A saida dos iões K+ também se faz de forma passiva pois a concentração destes iões é maior no interior das células ciliadas que no exterior do seu corpo celular que é banhado por perilinfa. Todo este processo resulta numa economia importante de ATP para as células ciliadas.
Estria vascular
A estrutura que sintetiza a endolinfa é a estria vascular. A estria vascular é uma estrutura epitelial complexa composta de vários tipos de células. As células basais e marginais são verdadeiras células epiteliais enquanto que as intermediárias são "melanócitos-like". Os vasos sanguíneos permitem o aporte de oxigénio e nutrientes necessário ao bom funcionamento do conjunto das células da estria vascular.

Estrutura da estria vascular (microscopia eletrónica de transmissão)
A estria vascular é ricamente vascularizada (C) e é constituida por 3 tipos de células:
as células marginais (M), que bordejam o canal cóclear (endolinfático) e que desencadeiam um papel fundamental na troca de iões.
as células intermediárias (I), ricas em pigmento melânico,
- as células basais (B).


Esta tarefa é executada pela Na,K-ATPase expressa nas membranas celulares das células intermedìárias e marginais. Para além disso, o co-transportador NKCC1, expresso na membrana das células marginais, intervém também nessa regulação utilizado o gradiente de sódio gerado pela Na,K-ATPase para fazr reentrar potássio na célula marginal. Este mecanismo de entrada de potássio nas células marginais é muito rentável pois utilizado uma única molécula de ATP é capaz de fazer entrar 5 iões de potássio (2 graças à K-ATPase e 3 graças à NHCC1). Por fim, os iões cloro, que entram ao mesmo tempo que os iões sódio e potássio, através do co-transportador NKCC1, são expulsos pelos canais de cloro CLC-K associados à sua subunidade reguladora, a bartina. As mutações da bartina são responsáveis pelo sindroma de Bartter, que associa a surdez com tubulopatia.
O potássio irá ser finalmente segregado para a rampa média da cóclea por canais potássicos KCNQ1 associados à sua unidade reguladora KCNE1. As mutações dos seus dois génes são responsáveis pelo sindroma de Jervell e Lange-Nielsen (Sindroma do QT longo), que associam surdez bilateral a alongamento do espaço QT cardíaco.
Facebook Twitter Google+